 生物大分子學習的教育門戶
生物大分子學習的教育門戶 
267: 血管內皮生長因子和血管新生(Vascular Endothelial Growth Factor (VegF) and Angiogenesis)
本文由Ethan Cartagena,Mariam Gelashvili,Jasmine Keyes和Elizabeth Rosenzweig撰寫和插圖,是羅格斯定量生物醫學研究所(Rutgers Institute for Quantitative Biomedicine)為本科生和研究生舉辦的為期一周的訓練營的一部分。該文章是2022-2023年PDB-101健康重點"癌症生物學和治療學"(Cancer Biology and Therapeutics)中的一篇。
創造新的血管
我們的身體不斷地創造新的血管,藉此將必要的營養分配給所有細胞。癌細胞(cancer cell)利用這個被稱為血管生成的過程來生長和轉移。血管生成是由信號蛋白血管內皮生長因子(vascular endothelial growth factor,VegF)刺激的,當它釋放到血液中時,啟動了一個複雜的分子舞蹈,從而在細胞之間形成開口。 前體內皮細胞(precursor endothelial cell)可以遷移到這些開口處並創建新血管的內壁。癌細胞,特別是具有最高轉移能力的最具侵略性的類型,操縱著血管生成的過程。透過誘導腫瘤中新血管的生長,癌症可以轉移其他器官的養分,並在周圍組織中創造出快速生長和擴散的途徑。
配對激活
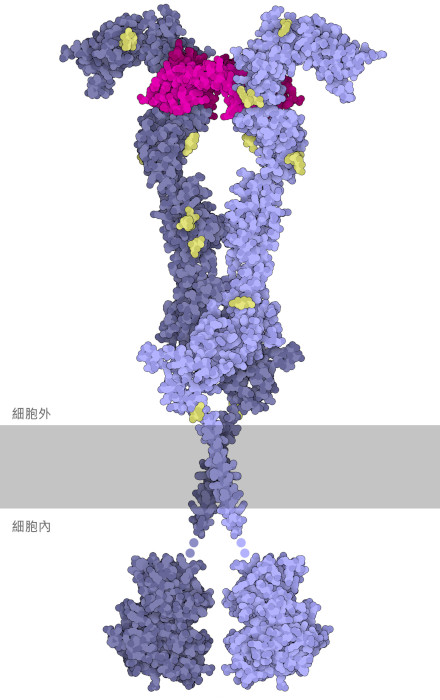
VegF攜帶啟動血管生成的訊號,這些訊號由VegF細胞受體上的一種稱為VegF受體酪胺酸激酶(VegF Receptor tyrosine kinase,VegFR)的跨膜蛋白接收。當處於非活性狀態時,受體作為一個單一的單元存在,像漂浮在水中的冰山一樣沿著細胞膜移動。然而,當它與VegF 結合時,它會與自身的另一個副本配對,形成活性二聚體,如圖所示。由於受體非常靈活且結構複雜,因此需要三個PDB 檔案來建立此插圖:胞外域(extracellular ectodomain,PDB條目5t89)、跨膜域(transmembrane domain,(PDB條目2m59),胞內酪胺酸激酶結構域(intracellular tyrosine kinase domain,PDB條目3hng)。二聚化過程將細胞膜內酪胺酸激酶結構域的兩個副本聚集在一起,使它們能夠相互活化。活性酪胺酸激酶反過來刺激細胞內的其他訊號蛋白。這開始了血管生成所需的許多過程,包括解開黏附結點(adherens junction),如下所述。
貝伐珠單抗:一把雙面刃
VegF的訊號傳遞是癌症生長的重要一步,它就成為癌症治療的一個有吸引力的標靶。 貝伐珠單抗(Bevacizumab,商品名阿瓦斯丁 Avastin)是一種單株抗體(monoclonal antibody),能與VegF結合並阻止其與受體結合。當VegF 透過與貝伐珠單抗結合而失去活性時,VegFR 二聚化的速度減慢,進而減慢血管生成的過程。這可以防止氧氣和營養物質過快到達腫瘤,並減少癌細胞離開原來的腫瘤並遷移到身體其他部位的機會。然而,雖然使用貝伐單抗治療能有效抑制癌細胞的生長,但它也會對其他過程造成嚴重的副作用,例如減緩傷口癒合。
鋪設管道
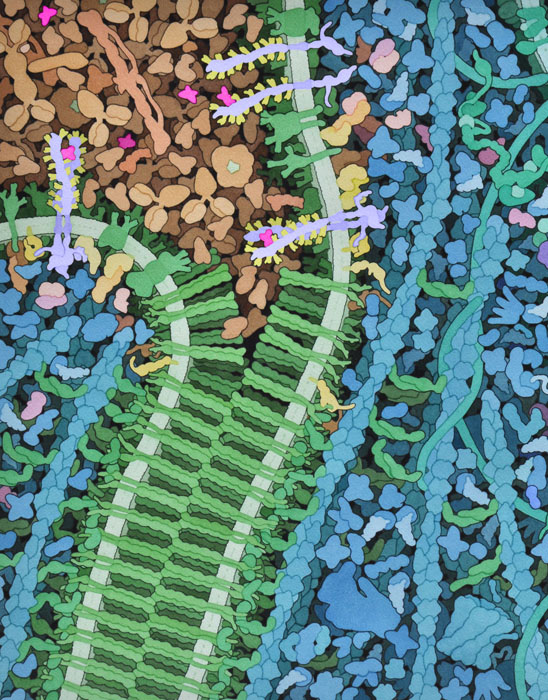
VegF的訊號傳遞是在營養缺乏的條件下開始的,例如快速生長的腫瘤所產生的條件,其中缺氧會刺激VegF訊號蛋白的產生和釋放,這些蛋白質通過血漿遷移並與VegFR的外域結合和激活。訊號被"傳送"過膜,VegFR的激酶域活化了細胞內的各種其他訊號傳導蛋白。其中一些轉移到細胞核並改變參與血管生成的基因的表達。訊號蛋白還能使細胞邊界的鈣粘蛋白(cadherin)的內部部分磷酸化。這促使鈣粘蛋白膜的外部部分打開,就像把尼龍搭扣撕成兩半一樣。前驅血管細胞被吸引到開口處並形成新血管的內壁。
傷口癒合
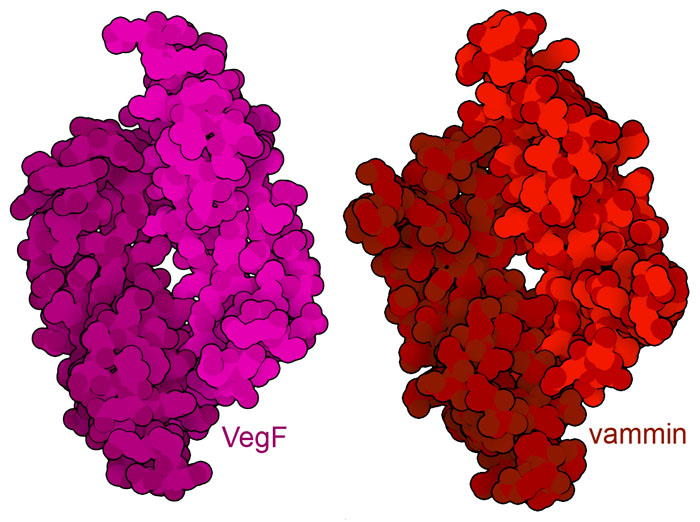
血管新生是控制轉移性腫瘤進展的重要標靶。然而,在其他情況下,我們可能想要刺激血管生成,以促進傷口癒合等有益過程。研究人員目前正在探索使用模仿VegF 結構的蛋白質的可能性,從而複製其功能。例如,一種蛋白質vammin(PDB條目1wq8)來自蛇毒,在三維結構和序列上都與VegF(PDB條目1vpf)非常相似,並且與VegF一樣,能活化VegF受體。
探索結構
若要切換到有互動控制的頁面,請點選圖表下方的按鈕。如果載入沒有開始,請嘗試點擊圖表。
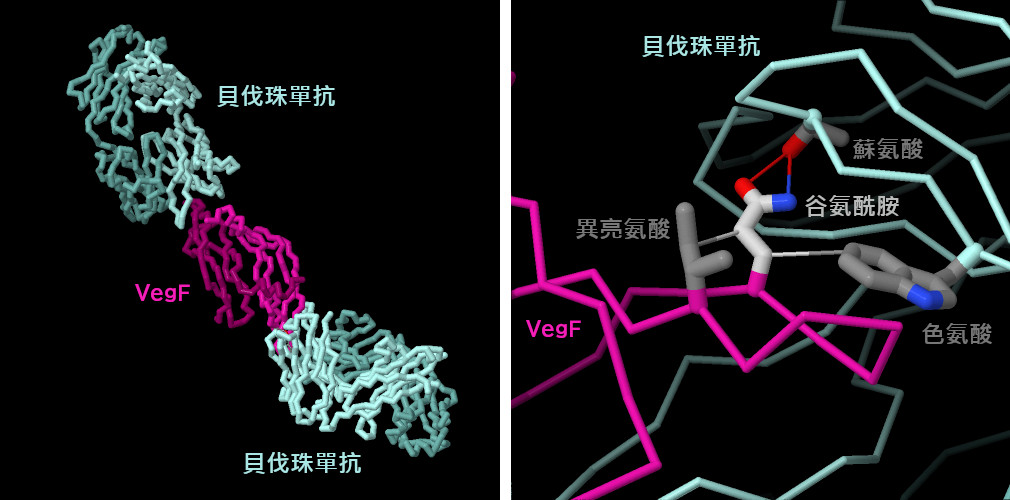
貝伐單抗與VegF的結構(PDB條目1bj1)顯示,兩個貝伐單抗分子與VegF相互作用,抑制VegF二聚體的兩端(圖的左側)。因此,此受體無法二聚並活化一系列與血管生成有關的訊號。與大多數抗體一樣,貝伐珠單抗和VegF之間形成高度互補的相互作用。例如,以麩醯胺酸為中心的一組三種分子相互作用對於貝伐珠單抗與VegF的結合至關重要。這些包括與相鄰的蘇氨酸的氫鍵相互作用,以及與色氨酸和異亮氨酸的疏水作用,這些相互作用將谷氨酰胺限制在正確的構像中。點擊圖下方的按鈕,切換到可操作的互動式圖像,以更詳細地了解這個結構。
進一步的討論議題
參考文獻
- 2021 A structural overview of vascular endothelial growth factors pharmacological ligands: from macromolecules to designed peptidomimetics. Molecules 26 6759
- 5t89 2017 Structure of the full-length VEGFR-1 extracellular domain in complex with VEGF-A. Structure 25 341-352
- 2017 Snake venom VEGF Vammin induces a highly efficient angiogenic response in skeletal muscle via VEGFR-2/NRP specific signaling. Scientific reports 7 5525
- 2015 Targeting angiogenesis in cancer therapy: moving beyond vascular endothelial growth factor. Oncologist 20 660-673
- 2m59 2014 Structural and functional characterization of alternative transmembrane domain conformations in VEGF receptor 2 activation. Structure 22 1077-1089
- 2013 Protein structure in context: the molecular landscape of angiogenesis. Biochemistry and Molecular. Biology Education 41 213-223
- 2010 Bevacizumab. Oncologist 15 819-825
- 1wq8 2005 Crystal structures of novel vascular endothelial growth factors (VEGF) from snake venoms: insight into selective VEGF binding to kinase insert domain-containing receptor but not to fms-like tyrosine kinase-1. Journal of Biological Chemistry 280 2126-2131
- 1bj1 1998 VEGF and the Fab fragment of a humanized neutralizing antibody: crystal structure of the complex at 2.4 Å resolution and mutational analysis of the interface. Structure 6 1153-1167
- 1vpf 1997 Vascular endothelial growth factor: crystal structure and functional mapping of the kinase domain receptor binding site. Proceedings of the National Academy of Sciences 94 7192-7197