 の生体高分子学習ポータルサイト
の生体高分子学習ポータルサイト 
248: フィトスルホカイン受容体(Phytosulfokine Receptor)
植物は非常に強靭である。私は庭でタンポポを取り除こうとしているときにいつもこう思う。バラなどの植物は、枝を取って地面に挿し世話をしてやれば健全な新しい植物となるだろう。植物細胞は自身に何が起こっているのかに基づいて常に決定を下している。いつ成長しどのようにして傷を治すのかを決め、どの部分が根となりどの部分が花となるのかを判断している。また、オーキシン(auxin)のような低分子やフィトスルホカイン(phytosulfokine)のようなペプチドなどを含む何百種類ものホルモンを介して隣接する細胞と常に通信を行い、これらの決定を下している。
オンにするスイッチ
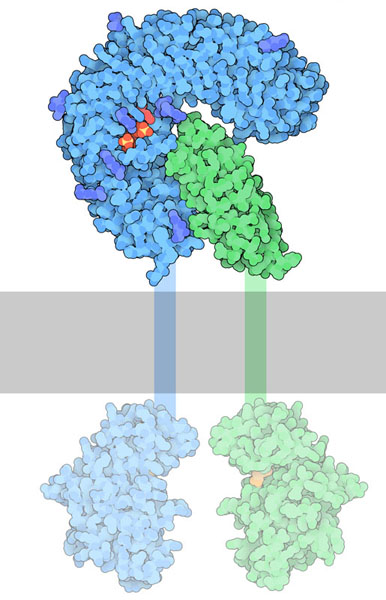
ホルモンの一つフィトスルホカインは「さぁ、成長しなさい」という単純なメッセージを送る。これは5つのアミノ酸でできた小さなペプチドで、細胞から分泌され、細胞表面にある膜結合受容体によって検知される。フィトスルホカインが受容体に結合すると、補助受容体タンパク質SERK1が急速に結合する。その様子はPDBエントリー4z64で確認できる。これにより、細胞内領域にあるキナーゼドメイン(kinase domain)が活性化され、細胞の成長を促す信号が始まる。キナーゼドメインは似た受容体(PDBエントリー5lpb)の構造を使って示している。
修飾された信号
植物細胞は何百種類もの低分子ペプチドホルモンをつくりさまざまなメッセージを送る。例えば、システミン(systemin)は植物が昆虫によって被害を受けたときの治療管理に、CLAVATAホルモンは急速に成長する根や芽の先端の発達制御に役立つ。通常これらのホルモンは、まずより大きなタンパク質としてつくられ、その後これに特化したプロテアーゼによって適切な大きさへと切断される。またペプチドは、糖の付加やプロリンへのヒドロキシ基付加などの化学修飾によってさらに調整されていることもよくある。フィトスルホカインは名前から想像されるように、2つのチロシンアミノ酸に硫酸基(sulfate group)が付加されている。
環形で独特なロイシンリピート
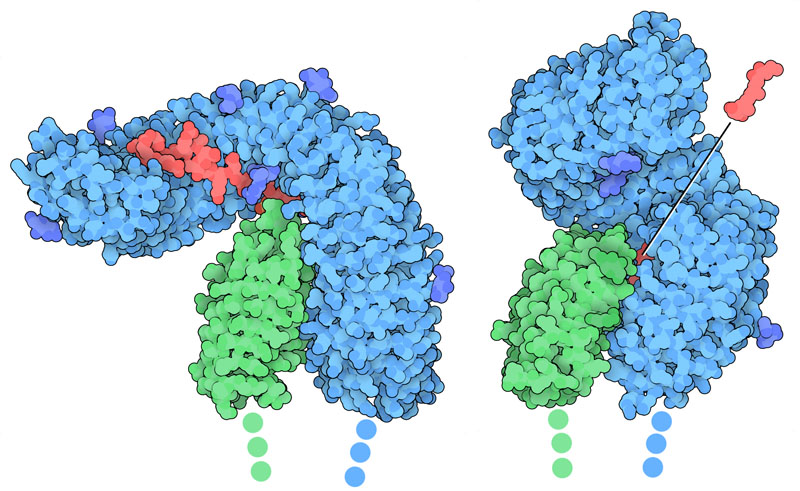
植物のホルモン受容体は、その多くが特有の曲がった形をとり、ロイシンリッチリピート(leucine-rich repeat、ロイシンが多くみられる繰り返し)配列がたくさん並んでできている。ここには似た受容体を2つ示す。これらは結合部位の構築にロイシンリピートが用いられている。FLS2(フラジェリン感受性2、flagellin-sensitive 2)は植物の免疫系に関係している(PDBエントリー4mn8)。FLS2は細菌が持つタンパク質の一種フラジェリン(flagellin)を認識し、防護反応を発動する。このとき補助受容体BAK1の助けを借りる。BRI1(ブラシノステロイド非感受性1、brassinosteroid insensitive 1)はステロイド植物ホルモンを認識し、補助受容体SERK1の助けを借りて成長を制御する(PDBエントリー4lsx)。
構造をみる
対話的操作のできるページに切り替えるには図の下のボタンをクリックしてください。読み込みが始まらない時は図をクリックしてみてください。
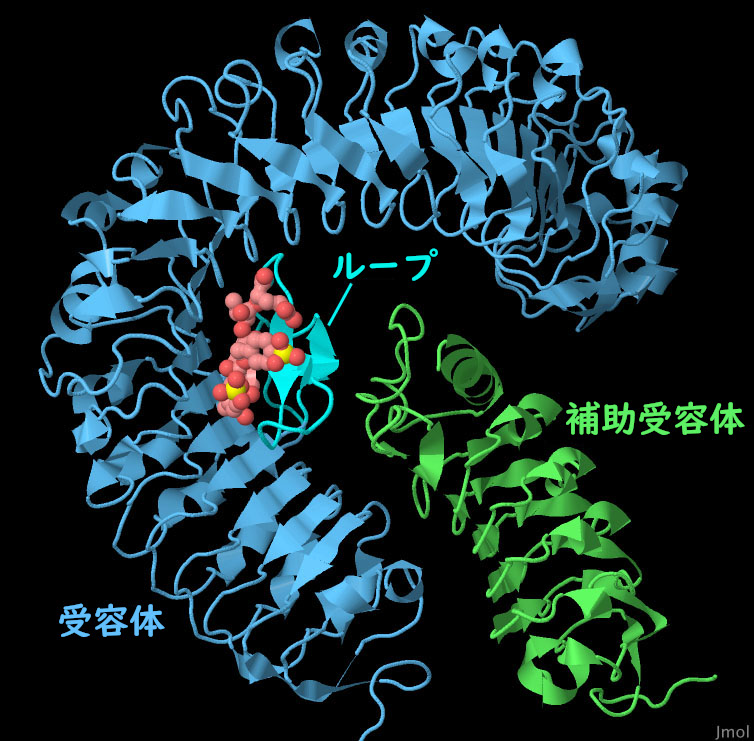
上に示すFLS2では、フラジェリンが糊のような役割を果たし、受容体と補助受容体をくっつけている。一方、フィトスルホカイン受容体ではこれを2段階かけて行う。フィトスルホカインが結合することにより、受容体の柔軟なループ領域が折りたたまれてホルモンを捕らえる安定な窪みを形成する。この複合体は補助受容体が結合する上で申し分のない形になっている。3つの構造がこの過程を示している。何も結合していない受容体(PDBエントリー4z62)ではループ領域の柔軟性が高すぎ構造中では見えていない。PDBエントリー4z63の構造はペプチドと受容体の複合体を示している。PDBエントリー4z64は最終的な活性化された複合体を示している。これらの構造をより詳しくみるため、図の下のボタンをクリックして対話的操作のできる画像に切り替えてみて欲しい。
理解を深めるためのトピックス
- ロイシンリッチリピートでできたタンパク質は興味深い機能を持っている。PDBjのウェブサイトでロイシンリッチリピートのキーワードを使って検索し、それらを見てみよう。
- これらの受容体は通常細胞外部分の構造しか含まれていない。RCSB PDBのProtein Explorerでどの部分が構造に含まれていないのかを確認してみよう。
参考文献
- 2015 Phytosulfokine peptide signalling. J. Exper. Botany 66 5161-5169
- 4z62, 4z63, 4z64 2015 Allosteric receptor activation by the plant peptide hormone phytosulfokine. Nature 525 265-268
- 2014 Posttranslationally modified small-peptide signals in plants. Annu. Rev. Plant Biol. 65 385-413
- 4lsx 2013 Molecular mechanism for plant steroid receptor activation by somatic embryogenesis co-receptor kinases. Science 341 889-892
- 4mn8 2013 Structural basis for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science 342 624-628