 の生体高分子学習ポータルサイト
の生体高分子学習ポータルサイト 
238: リボヌクレオチド還元酵素(Ribonucleotide Reductase)
DNAとRNAはほとんど同じ構造をしているが、一つの小さな違いが大きな結果の違いをもたらしている。DNAのヌクレオチドは酸素が一つ欠けているが、これがDNAとRNAの違いになっている。小さな違いだが、この酸素がないことでDNAはより安定となり、情報を長期間保持するのに適した分子になっている。一方、RNAはDNAに比べると不安定である。RNAだけが持つ追加の酸素はヒドロキシル基をつくり、RNAをより加水分解されやすくしている。だからRNAは、一時的な生物学的データを保持し必要なくなると分解される、生物の「フラッシュドライブ」のような働きをすることがよくある。私たちの体内でデオキシヌクレオチドをつくる唯一の方法は、酵素のリボヌクレオチド還元酵素(ribonucleotide reductase)を使いリボヌクレオチドからつくり出すという方法である。これはDNAの合成と修復にとって欠かせない。
ラジカルを使う過激な酵素
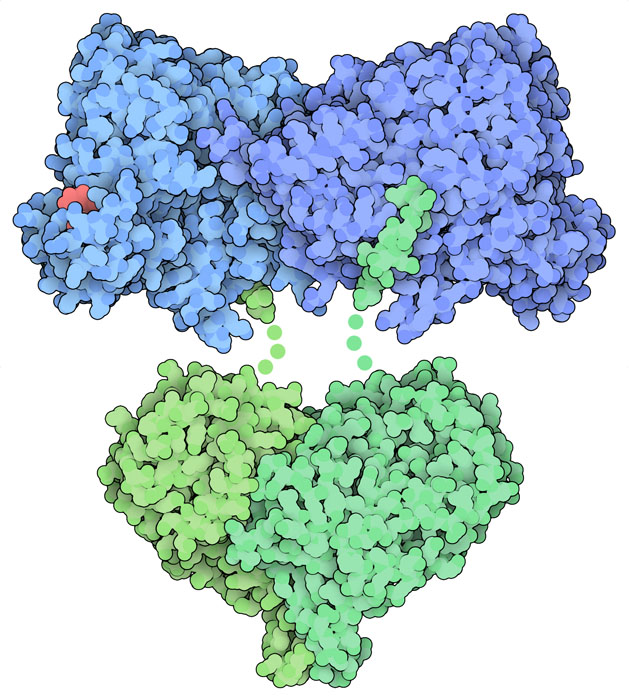
リボヌクレオチド還元酵素による触媒のしくみは興味深い。なぜならフリーラジカル(free radical、遊離基)を必要とするからである。通常私たちの体にとってフリーラジカルは有害であるが、DNAの構成部材をつくる際にリボヌクレオチド還元酵素の中で不可欠な役割を果たす。I型酵素では(R2とも呼ばれる)βサブユニットでフリーラジカルが生成され、それがより大きな触媒サブユニットとなるαサブユニット(R1)まで運ばれ、ここでデオキシリボヌクレオチド(deoxyribonucleotide)がつくられる。リボヌクレオチド還元酵素はあらゆる生命においてよく保存された酵素だが、この必要不可欠なフリーラジカルを生成する際に用いる金属が違うさまざまな種類のものがある。ここに示すのはよく研究された細菌由来のリボヌクレオチド還元酵素が持つ活性部位の構造(PDBエントリー1mrr、3r1r)で、各サブユニットを2つずつ持ちそれらは柔軟な接続部でつながっている。
がん治療の対象
リボヌクレオチド還元酵素は抗がん剤にとって重要な作用対象となる。がん細胞の成長を止める一つの方法は、DNA合成に関わる酵素を止めることである。この酵素を阻害するという明確な方法は、ヌクレオシドに似ていてしかも活性部位で通常ならヌクレオシド2リン酸が結合するところを阻害する分子をつくる、というやり方になるだろう。しかし、ヌクレオシド類似物質は細胞内で実際に使われるヌクレオシドと非常によく似ているので、がん細胞だけではなく健康な細胞においてもDNAに組み込まれ副作用を起こしてしまう。この問題を改善する一つの方法は、ヌクレオシド類似物質ではないがリボヌクレオチド還元酵素は選択的に阻害する小さな分子をつくるという方法である。両方の研究手法を使った事例を構造をみるの節で示している。
チェックとバランス
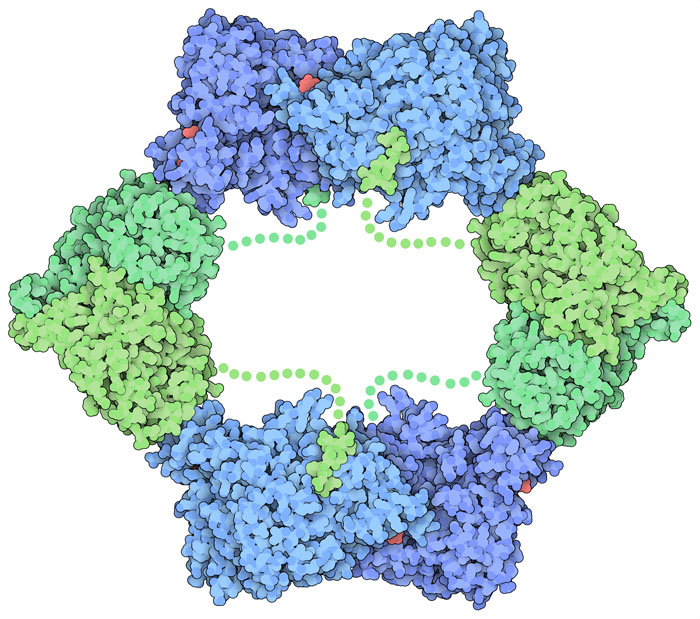
リボヌクレオチド還元酵素はDNA合成にとって不可欠であるため、細胞ではしっかり制御され必要なときにだけ働くようになっている。細胞内のATP濃度が高くなると、ATPはリボヌクレオチド還元酵素に結合し、それが信号となってより多くのデオキシリボヌクレオチドがつくられる。これは通常、大量の新たなDNAが一気につくられる細胞分裂のときに起こることである。さらに、リボヌクレオチド還元酵素にはデオキシリボヌクレオチドがが必要とされていることを検知する別の部位もある。酵素が有害な濃度のデオキシリボヌクレオチドをつくってしまわないようにするため、dATPは酵素に結合して働きを止める。酵素の働きを止める方法は非常に巧妙で、PDBエントリー3uusで見られるように酵素が持つ複数のサブユニットが互いに結合し、環状の構造をつくる。この分子再配置により、フリーラジカルがβサブユニットからαサブユニットへと移動する経路が遮断される。
構造をみる
対話的操作のできるページに切り替えるには図の下のボタンをクリックしてください。読み込みが始まらない時は図をクリックしてみてください。
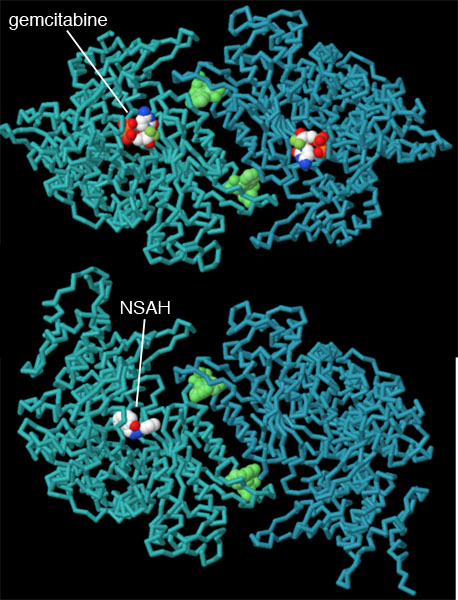
2つの合成分子によって阻害されたリボヌクレオチド還元酵素の構造(PDBエントリー5tus、2eud)をここに示す。ゲムシタビン(gemcitabine)は現在さまざまな種類のがんを治療するのに使われるヌクレオシド類似物質の抗がんプロドラッグ(体内で代謝されて薬効のある抗がん剤に変化する物質)である。ゲムシタビンは触媒部位に結合し、通常のヌクレオシドであれば水素原子とヒドロキシル基がある場所にフッ素原子を置いて酵素を「だます」ことによりαサブユニットの働きを阻害する。ナフチルサリチル酸ヒドラゾン(naphthyl salicylic acyl hydrazone、NSAH)はin silicoスクリーニング(コンピュータを使った化合物検索)によって発見された非ヌクレオチド阻害剤である。この化合物も触媒部位に結合し、リボヌクレオチド還元酵素の働きを可逆的に阻害する。図の下のボタンを押して対話的操作のできる画像に切り替えると、これらの構造をより詳しく比較することができる。
理解を深めるためのトピックス
- PDBアーカイブにはDNAとRNAの両方が含まれた構造がたくさんある。相違点となっているRNA中の糖にあるヒドロキシル基を見てみて欲しい。PDBエントリー1g4qから始めると分かりやすいだろう。この構造には1本がDNAでもう1本がRNAになっている混合型二重らせんの短いものが含まれている。
参考文献
- 5tus 2017 Potent competitive inhibition of human ribonucleotide reductase by a nonnucleoside small molecule. Proc. Natl. Acad. Sci. U.S.A. 114 8241-8246
- 3uus 2011 Structural interconversions modulate activity of Escherichia coli ribonucleotide reductase. Proc.Natl.Acad.Sci.USA 108 21046-21051
- 2eud 2006 Structures of eukaryotic ribonucleotide reductase I define gemcitabine diphosphate binding and subunit assembly. Proc.Natl.Acad.Sci.Usa 103 4028-4033
- 3r1r 1997 Binding of allosteric effectors to ribonucleotide reductase protein R1: reduction of active-site cysteines promotes substrate binding. Structure 5 1077-1092
- 1mrr 1992 Substitution of manganese for iron in ribonucleotide reductase from Escherichia coli. Spectroscopic and crystallographic characterization. J.Biol.Chem. 267 20682-20688